🇦🇹🇬🇧🇦🇹🇬🇧🇦🇹🇬🇧🇦🇹

(Gracillariidae: leaf-mining larvae and host plant coevolution)
Introduzione
Italiano
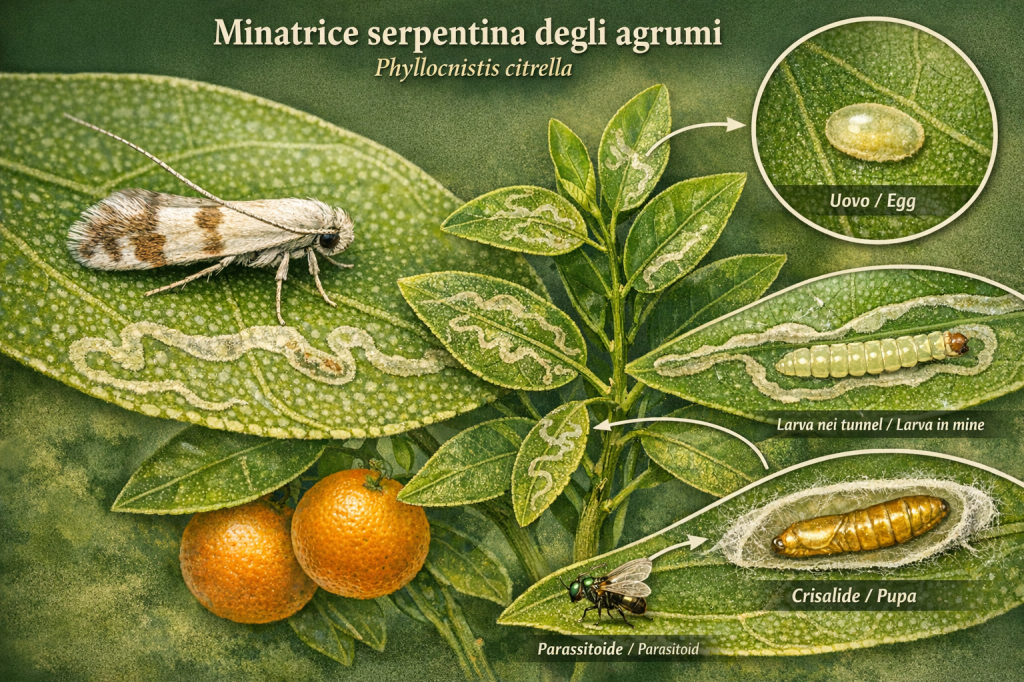
I Gracillariidae rappresentano una famiglia di microlepidotteri altamente specializzati, caratterizzati da larve mineratrici che si sviluppano all’interno dei tessuti fogliari delle piante ospiti. Questa famiglia comprende generi come Phyllonorycter e Caloptilia, noti per la loro stretta specificità dell’ospite e per il loro ruolo ecologico nei sistemi agricoli e naturali. La loro presenza evidenzia come piccole specie possano avere un impatto significativo sulla fisiologia vegetale e sulle dinamiche ecologiche dei microhabitat.
L’osservazione e lo studio dei Gracillariidae offrono una finestra sulla coevoluzione tra insetti e piante, mostrando adattamenti larvali unici per sfruttare risorse interne protette delle foglie e strategie di sopravvivenza contro predatori e parassitoidi.
English
Gracillariidae represent a family of highly specialized microlepidopterans, characterized by leaf-mining larvae that develop within the leaf tissues of their host plants. This family includes genera such as Phyllonorycter and Caloptilia, known for their host specificity and ecological roles in agricultural and natural systems. Their presence demonstrates how small species can significantly influence plant physiology and ecological dynamics within microhabitats.
Observation and study of Gracillariidae provide insight into insect-plant coevolution, highlighting unique larval adaptations for exploiting protected internal leaf resources and survival strategies against predators and parasitoids.
Morfologia e ciclo vitale
Italiano
Gli adulti dei Gracillariidae sono piccolissimi, con apertura alare di 5–10 mm, ali strette e spesso con riflessi metallici. Le femmine depongono le uova sulla superficie superiore o inferiore delle foglie giovani, preferendo tessuti in rapida crescita.
Le larve emergono e iniziano a scavare gallerie tra gli strati epidermici, nutrendosi dei tessuti fotosintetici. Alcune specie di Caloptilia completano inizialmente il ciclo come minatrici e successivamente passano a costruire pieghe fogliari o sacche protettive. La fase larvale dura generalmente 2–3 settimane, seguita dalla fase pupale, spesso protetta all’interno della foglia o in case di seta, prima della comparsa dell’adulto.
English
Adult Gracillariidae are very small, with a wingspan of 5–10 mm, narrow wings often exhibiting metallic reflections. Females lay eggs on the upper or lower surface of young leaves, preferring rapidly growing tissues.
The larvae hatch and begin creating mines between the epidermal layers, feeding on photosynthetic tissues. Some Caloptilia species initially develop as miners and later transition to folding leaves or building protective silk cases. The larval stage generally lasts 2–3 weeks, followed by pupation, often protected within the leaf or silk case, before adult emergence.
Impatto ecologico e agricolo
Italiano
Le attività minerarie delle larve possono ridurre la capacità fotosintetica delle foglie, causando stress vegetativo, deformazioni dei germogli e, in casi di infestazioni elevate, diminuzione della produttività delle piante coltivate. Tuttavia, le Gracillariidae svolgono anche un ruolo ecologico significativo come prede di parassitoidi e predatori, contribuendo a mantenere l’equilibrio degli ecosistemi agricoli e naturali.
Il monitoraggio delle popolazioni è fondamentale per la gestione sostenibile, integrando metodi biologici con pratiche agronomiche volte a preservare la salute delle coltivazioni.
English
Larval mining activity can reduce the photosynthetic capacity of leaves, causing vegetative stress, shoot deformation, and, in cases of severe infestation, decreased crop productivity. However, Gracillariidae also play a significant ecological role as prey for parasitoids and predators, helping maintain balance in agricultural and natural ecosystems.
Population monitoring is essential for sustainable management, integrating biological methods with agronomic practices aimed at preserving crop health.
Strategie di gestione
Italiano
Il controllo delle Gracillariidae richiede un approccio integrato. L’introduzione o la conservazione di parassitoidi naturali, come Hymenoptera piccoli e specialisti, è altamente efficace nel ridurre le larve. Le tecniche agronomiche includono la potatura dei germogli infestati e la gestione attenta dell’irrigazione e della fertilizzazione per migliorare la resistenza delle piante. L’uso di insetticidi deve essere ponderato e limitato, privilegiando prodotti selettivi a basso impatto ecologico.
English
Control of Gracillariidae requires an integrated approach. The introduction or conservation of natural parasitoids, such as small, specialist Hymenoptera, is highly effective in reducing larval populations. Agronomic techniques include pruning infested shoots and careful management of irrigation and fertilization to improve plant resistance. Pesticide use should be carefully considered and limited, favoring selective, low-impact products.
Conclusione
Italiano
I Gracillariidae rappresentano un esempio eccellente di microlepidotteri specialistici, la cui osservazione permette di comprendere la coevoluzione con le piante ospiti, le strategie di difesa larvale e l’interazione con l’ecosistema circostante. La conoscenza approfondita di queste specie è fondamentale per una gestione sostenibile delle coltivazioni e per lo studio della biodiversità entomologica.
English
Gracillariidae represent an excellent example of specialist microlepidopterans, whose study provides insights into coevolution with host plants, larval defense strategies, and interactions with surrounding ecosystems. In-depth knowledge of these species is essential for sustainable crop management and the study of entomological biodiversity.