Conservative Evolution in Lepidoptera: The Same Genes Used for 120 Million Years
Introduzione
L’evoluzione viene spesso immaginata come un processo caotico, imprevedibile e completamente casuale. Nella percezione comune ogni nuova forma biologica sarebbe il risultato di mutazioni casuali che, nel corso del tempo, producono organismi differenti. Tuttavia le moderne ricerche di genetica evolutiva mostrano una realtà molto più complessa. Numerosi studi sui lepidotteri, in particolare sulle farfalle tropicali caratterizzate da colorazioni spettacolari, dimostrano che l’evoluzione tende frequentemente a riutilizzare gli stessi strumenti genetici per milioni di anni. Alcuni geni coinvolti nello sviluppo delle ali e dei pattern cromatici sono rimasti attivi e conservati per oltre 120 milioni di anni, attraversando ere geologiche, estinzioni di massa, cambiamenti climatici globali e profonde trasformazioni ecologiche.
Questo fenomeno suggerisce che la selezione naturale non opera esclusivamente attraverso l’invenzione continua di nuove strutture genetiche, ma anche attraverso il riutilizzo di sistemi biologici estremamente efficienti. I geni antichi possono essere modificati, regolati o combinati in modi differenti, producendo una sorprendente varietà di forme senza la necessità di creare da zero nuove soluzioni biologiche.
La scoperta rappresenta uno dei punti più affascinanti della moderna biologia evolutiva, poiché collega genetica, ecologia, sviluppo embrionale e adattamento ambientale. Nel caso delle farfalle, i ricercatori hanno osservato che i medesimi geni responsabili dei colori delle ali in specie moderne risultano associati a strutture simili in antenati antichissimi.
Introduction
Evolution is often imagined as a chaotic, unpredictable, and completely random process. In common perception, every new biological form would emerge from random mutations that over time generate different organisms. However, modern research in evolutionary genetics reveals a far more complex reality. Numerous studies on lepidopterans, especially tropical butterflies with spectacular wing coloration, demonstrate that evolution frequently reuses the same genetic tools for millions of years. Some genes involved in wing development and color pattern formation have remained active and conserved for more than 120 million years, surviving geological eras, mass extinctions, global climate shifts, and profound ecological transformations.
This phenomenon suggests that natural selection does not operate exclusively through the constant invention of new genetic structures, but also through the reuse of highly efficient biological systems. Ancient genes can be modified, regulated, or recombined in different ways, producing extraordinary diversity without requiring entirely new biological inventions.
This discovery represents one of the most fascinating aspects of modern evolutionary biology because it connects genetics, ecology, embryonic development, and environmental adaptation. In butterflies, researchers observed that the same genes responsible for wing coloration in modern species are associated with similar structures in extremely ancient ancestors.
Le Origini Evolutive dei Lepidotteri
I lepidotteri costituiscono uno degli ordini di insetti più diversificati del pianeta. Comprendono farfalle diurne, falene notturne e numerose forme intermedie adattate ai più diversi ecosistemi terrestri. Le prime linee ancestrali comparvero probabilmente nel Giurassico, in un periodo dominato dai dinosauri e caratterizzato da immense foreste primitive.
L’origine delle ali squamose rappresentò una rivoluzione evolutiva. Le minuscole scaglie che ricoprono le ali dei lepidotteri non sono semplici strutture decorative, ma complessi organi microscopici in grado di riflettere, assorbire e diffondere la luce. Attraverso queste strutture gli insetti svilupparono strategie di mimetismo, comunicazione sessuale, difesa dai predatori e regolazione termica.
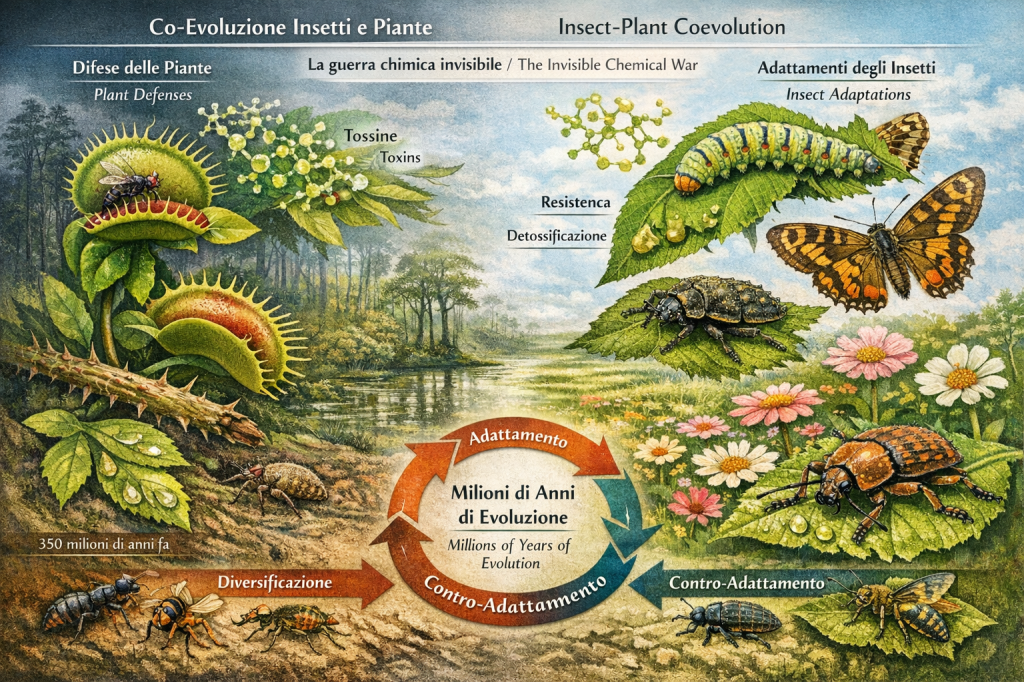
Nel corso di milioni di anni le farfalle si diversificarono insieme alle piante da fiore. Questo rapporto coevolutivo trasformò profondamente gli ecosistemi terrestri. Le piante svilupparono colori, profumi e nettare per attirare impollinatori, mentre le farfalle svilupparono apparati sensoriali sempre più sofisticati.
The Evolutionary Origins of Lepidoptera
Lepidoptera constitute one of the most diverse insect orders on Earth. They include butterflies, moths, and numerous intermediate forms adapted to highly different terrestrial ecosystems. The earliest ancestral lineages probably appeared during the Jurassic period, an era dominated by dinosaurs and immense primitive forests.
The origin of scaled wings represented an evolutionary revolution. The tiny scales covering butterfly wings are not merely decorative structures but complex microscopic organs capable of reflecting, absorbing, and scattering light. Through these structures insects developed strategies of camouflage, sexual communication, predator defense, and thermal regulation.
Over millions of years butterflies diversified alongside flowering plants. This coevolutionary relationship deeply transformed terrestrial ecosystems. Plants evolved colors, scents, and nectar to attract pollinators, while butterflies developed increasingly sophisticated sensory systems.
Conservazione Genetica e Stabilità Evolutiva
Uno degli aspetti più sorprendenti emersi dagli studi recenti riguarda la stabilità di determinati circuiti genetici. Per lungo tempo gli scienziati immaginarono che grandi differenze morfologiche derivassero principalmente dalla comparsa di geni completamente nuovi. Oggi sappiamo che spesso l’innovazione biologica nasce dalla regolazione differente di geni molto antichi.
I geni coinvolti nella pigmentazione delle ali delle farfalle appartengono frequentemente a famiglie genetiche conservate. Questi geni controllano processi fondamentali come la produzione di melanina, la disposizione delle squame e la formazione di pattern geometrici complessi.
La conservazione genetica non significa immobilità evolutiva. Al contrario, indica che alcune soluzioni biologiche sono così efficienti da essere mantenute per tempi geologici enormi. L’evoluzione lavora come un ingegnere che modifica progetti esistenti invece di costruire ogni volta un sistema completamente nuovo.
Genetic Conservation and Evolutionary Stability
One of the most surprising aspects emerging from recent studies concerns the stability of certain genetic circuits. For a long time scientists imagined that major morphological differences mainly resulted from the appearance of entirely new genes. Today we know that biological innovation often arises from the differential regulation of extremely ancient genes.
Genes involved in butterfly wing pigmentation frequently belong to conserved genetic families. These genes control fundamental processes such as melanin production, scale arrangement, and the formation of complex geometric patterns.
Genetic conservation does not mean evolutionary immobility. On the contrary, it indicates that some biological solutions are so efficient that they are maintained across enormous geological timescales. Evolution works like an engineer modifying existing blueprints rather than creating a completely new system every time.
Pattern Cromatici e Comunicazione Visiva
Le ali delle farfalle costituiscono una delle superfici biologiche più sofisticate del regno animale. I colori possono derivare da pigmenti chimici oppure da fenomeni strutturali legati alla rifrazione della luce. In molte specie tropicali le tonalità metalliche blu e verdi non dipendono da pigmenti ma dalla disposizione nanometrica delle squame.
I pattern cromatici svolgono funzioni multiple. Alcune specie imitano organismi tossici per scoraggiare i predatori. Altre utilizzano colori vivaci per il riconoscimento sessuale. In certi casi le ali imitano foglie secche, corteccia o superfici vegetali.
La genetica moderna ha dimostrato che pochi geni regolatori possono controllare enormi variazioni visive. Modificando leggermente l’attività di un gene durante lo sviluppo larvale è possibile alterare profondamente l’aspetto finale dell’insetto adulto.
Color Patterns and Visual Communication
Butterfly wings represent one of the most sophisticated biological surfaces in the animal kingdom. Colors may derive from chemical pigments or from structural phenomena linked to light refraction. In many tropical species metallic blue and green tones do not depend on pigments but on the nanometric arrangement of scales.
Color patterns serve multiple functions. Some species mimic toxic organisms to discourage predators. Others use bright colors for sexual recognition. In certain cases wings imitate dry leaves, bark, or plant surfaces.
Modern genetics has demonstrated that a few regulatory genes can control enormous visual variations. Slightly modifying the activity of a gene during larval development can profoundly alter the final appearance of the adult insect.
Il Gene Optix e la Colorazione Rossa
Uno dei geni più studiati nei lepidotteri è optix, associato alla formazione delle colorazioni rosse e arancioni nelle ali. Questo gene è presente in numerose specie evolutivamente distanti tra loro, suggerendo un’origine estremamente antica.
La funzione di optix mostra come l’evoluzione riutilizzi moduli genetici esistenti. Lo stesso gene può produrre effetti differenti a seconda del contesto cellulare e della combinazione con altri fattori regolatori.
Nelle farfalle tropicali del genere Heliconius il gene optix controlla pattern di mimetismo complessi che permettono a specie differenti di apparire quasi identiche. Questo fenomeno rappresenta uno straordinario esempio di convergenza evolutiva guidata da meccanismi genetici condivisi.
The Optix Gene and Red Coloration
One of the most studied genes in Lepidoptera is optix, associated with the formation of red and orange coloration on butterfly wings. This gene is present in numerous evolutionarily distant species, suggesting an extremely ancient origin.
The function of optix illustrates how evolution reuses existing genetic modules. The same gene can produce different effects depending on the cellular context and its interaction with other regulatory factors.
In tropical butterflies of the genus Heliconius, the optix gene controls complex mimicry patterns allowing different species to appear almost identical. This phenomenon represents an extraordinary example of evolutionary convergence driven by shared genetic mechanisms.
Selezione Naturale e Predazione
La pressione esercitata dai predatori costituisce uno dei principali motori dell’evoluzione delle ali delle farfalle. Gli uccelli, i rettili e persino altri insetti selezionano continuamente individui più difficili da individuare o meno appetibili.
Nel corso di milioni di anni questo processo ha favorito la comparsa di occhi finti, bande colorate, motivi geometrici e strategie mimetiche incredibilmente sofisticate. Alcune falene possiedono ali che imitano perfettamente il legno coperto di licheni. Altre simulano serpenti o occhi di vertebrati.
La sorprendente scoperta è che molti di questi adattamenti derivano dalla riattivazione di antichi circuiti genetici piuttosto che dalla nascita di geni completamente nuovi.
Natural Selection and Predation
Predatory pressure represents one of the primary driving forces behind butterfly wing evolution. Birds, reptiles, and even other insects continuously select individuals that are harder to detect or less palatable.
Over millions of years this process favored the emergence of false eyes, colored bands, geometric motifs, and highly sophisticated mimicry strategies. Some moths possess wings perfectly imitating lichen-covered wood. Others simulate snakes or vertebrate eyes.
The remarkable discovery is that many of these adaptations arise from the reactivation of ancient genetic circuits rather than from the birth of entirely new genes.
Evo-Devo e Biologia dello Sviluppo
La disciplina conosciuta come evo-devo, abbreviazione di evolutionary developmental biology, ha rivoluzionato la comprensione dell’evoluzione animale. Gli studiosi hanno compreso che piccoli cambiamenti nei tempi e nei luoghi di attivazione dei geni possono produrre enormi differenze morfologiche.
Durante lo sviluppo della crisalide le cellule delle ali ricevono segnali biochimici estremamente precisi. Questi segnali determinano la disposizione delle scaglie, la loro forma e la pigmentazione finale.
L’evo-devo dimostra che organismi molto differenti condividono strumenti genetici comuni. Geni simili controllano lo sviluppo degli occhi negli insetti e nei vertebrati, evidenziando una profonda unità evolutiva della vita.
Evo-Devo and Developmental Biology
The discipline known as evo-devo, short for evolutionary developmental biology, revolutionized our understanding of animal evolution. Researchers realized that small changes in the timing and location of gene activation can produce enormous morphological differences.
During chrysalis development, wing cells receive extremely precise biochemical signals. These signals determine scale arrangement, shape, and final pigmentation.
Evo-devo demonstrates that highly different organisms share common genetic tools. Similar genes control eye development in insects and vertebrates, revealing a deep evolutionary unity of life.
Le Farfalle come Modello Scientifico
Le farfalle rappresentano organismi modello ideali per lo studio dell’evoluzione. I loro cicli vitali relativamente brevi, la grande varietà di pattern cromatici e la facilità di osservazione permettono agli scienziati di analizzare rapidamente cambiamenti genetici e adattamenti ecologici.
Le collezioni museali hanno avuto un ruolo fondamentale. Attraverso campioni conservati per secoli i ricercatori possono confrontare DNA antico e moderno, ricostruendo la storia evolutiva delle popolazioni.
L’utilizzo delle moderne tecnologie di sequenziamento genetico ha reso possibile identificare reti genetiche condivise tra specie separate da decine di milioni di anni.
Butterflies as Scientific Models
Butterflies represent ideal model organisms for the study of evolution. Their relatively short life cycles, enormous diversity of color patterns, and ease of observation allow scientists to rapidly analyze genetic changes and ecological adaptations.
Museum collections played a fundamental role. Through specimens preserved for centuries, researchers can compare ancient and modern DNA, reconstructing the evolutionary history of populations.
The use of modern genetic sequencing technologies made it possible to identify genetic networks shared by species separated by tens of millions of years.
Coevoluzione con le Piante
L’evoluzione delle farfalle non può essere separata dall’evoluzione delle piante ospiti. Le larve dipendono da specifiche specie vegetali per l’alimentazione, mentre gli adulti utilizzano il nettare come principale fonte energetica.
Questa relazione ha generato una corsa evolutiva continua. Le piante sviluppano tossine chimiche per difendersi dagli erbivori, mentre le farfalle evolvono meccanismi di detossificazione.
In alcune specie tropicali le larve accumulano sostanze tossiche provenienti dalle piante e le mantengono anche da adulte, diventando pericolose per i predatori.
Coevolution with Plants
Butterfly evolution cannot be separated from the evolution of host plants. Larvae depend on specific plant species for feeding, while adults use nectar as their primary energy source.
This relationship generated a continuous evolutionary arms race. Plants evolve chemical toxins to defend themselves from herbivores, while butterflies evolve detoxification mechanisms.
In some tropical species larvae accumulate toxic substances from plants and retain them into adulthood, becoming dangerous to predators.
Mimicry e Adattamento
Il mimetismo costituisce una delle strategie evolutive più spettacolari osservabili nei lepidotteri. Attraverso il mimetismo batesiano specie innocue imitano specie tossiche. Nel mimetismo mülleriano invece più specie tossiche convergono verso pattern simili per rafforzare il segnale di pericolo.
La genetica del mimetismo dimostra ancora una volta il ruolo centrale dei geni conservati. In numerosi casi le stesse regioni genomiche controllano pattern analoghi in specie differenti.
Questa scoperta suggerisce che l’evoluzione possieda una sorta di memoria biologica, nella quale soluzioni efficaci vengono mantenute e riutilizzate nel tempo.
Mimicry and Adaptation
Mimicry represents one of the most spectacular evolutionary strategies observable in Lepidoptera. Through Batesian mimicry harmless species imitate toxic species. In Müllerian mimicry multiple toxic species converge toward similar patterns to strengthen warning signals.
The genetics of mimicry once again demonstrates the central role of conserved genes. In many cases the same genomic regions control analogous patterns in different species.
This discovery suggests that evolution possesses a form of biological memory in which effective solutions are maintained and reused over time.
Implicazioni Filosofiche dell’Evoluzione Conservativa
La scoperta che gli stessi geni possano essere utilizzati per oltre 120 milioni di anni modifica profondamente il modo in cui interpretiamo l’evoluzione. La natura non appare più come un sistema puramente casuale, ma come un processo capace di conservare strutture efficienti attraverso epoche immense.
Questo non significa che l’evoluzione abbia uno scopo prestabilito. Significa però che la selezione naturale tende a mantenere soluzioni funzionali e a riadattarle continuamente.
La stabilità genetica evidenzia anche l’incredibile robustezza dei sistemi biologici. Alcuni meccanismi molecolari sono talmente ottimizzati da sopravvivere quasi invariati per tempi superiori all’esistenza di intere catene montuose.
Philosophical Implications of Conservative Evolution
The discovery that the same genes can be used for more than 120 million years profoundly changes how we interpret evolution. Nature no longer appears as a purely random system but as a process capable of preserving efficient structures across immense spans of time.
This does not mean that evolution has a predetermined goal. It does mean, however, that natural selection tends to preserve functional solutions and continuously readapt them.
Genetic stability also highlights the extraordinary robustness of biological systems. Some molecular mechanisms are so optimized that they survive almost unchanged for periods longer than the existence of entire mountain ranges.
Cambiamenti Climatici e Sopravvivenza
Nel corso della loro storia evolutiva le farfalle hanno attraversato periodi climatici estremi. Hanno vissuto ere calde tropicali, glaciazioni e drastici cambiamenti ambientali.
La capacità di riutilizzare antichi circuiti genetici potrebbe aver favorito la loro resilienza. Gli organismi che possiedono sistemi regolatori flessibili possono adattarsi più rapidamente alle nuove condizioni ambientali.
Oggi i cambiamenti climatici rappresentano una nuova sfida globale. Molte specie di lepidotteri stanno modificando distribuzione geografica, periodi riproduttivi e interazioni ecologiche.
Climate Change and Survival
Throughout their evolutionary history butterflies survived extreme climatic periods. They experienced warm tropical eras, ice ages, and drastic environmental transformations.
The ability to reuse ancient genetic circuits may have favored their resilience. Organisms possessing flexible regulatory systems can adapt more rapidly to new environmental conditions.
Today climate change represents a new global challenge. Many lepidopteran species are altering geographic distribution, reproductive timing, and ecological interactions.
Tecnologie Genomiche Moderne
Le moderne tecniche di sequenziamento hanno rivoluzionato l’entomologia. Gli scienziati possono oggi analizzare interi genomi in tempi estremamente ridotti rispetto al passato.
Questi strumenti permettono di identificare geni condivisi tra specie lontane e di ricostruire alberi evolutivi con precisione straordinaria.
L’integrazione tra genomica, microscopia elettronica e intelligenza artificiale sta aprendo nuove frontiere nella comprensione dell’evoluzione biologica.
Modern Genomic Technologies
Modern sequencing techniques revolutionized entomology. Scientists can now analyze entire genomes in dramatically shorter times compared to the past.
These tools make it possible to identify genes shared between distant species and reconstruct evolutionary trees with extraordinary precision.
The integration of genomics, electron microscopy, and artificial intelligence is opening new frontiers in the understanding of biological evolution.
Conclusione
Lo studio dei geni conservati nelle farfalle dimostra che l’evoluzione è un processo molto più sofisticato di quanto immaginato in passato. La natura non crea continuamente da zero, ma modifica, ricombina e perfeziona strutture genetiche antichissime.
Il fatto che alcuni geni siano rimasti funzionali per oltre 120 milioni di anni rappresenta una testimonianza straordinaria della continuità della vita sulla Terra. Attraverso guerre biologiche, estinzioni e trasformazioni climatiche, determinati meccanismi molecolari hanno continuato a plasmare le ali delle farfalle.
Le farfalle diventano così simboli viventi della memoria evolutiva del pianeta. Ogni loro ala contiene tracce di un passato remoto che risale all’epoca dei dinosauri.
La ricerca futura probabilmente rivelerà che fenomeni simili esistono in molti altri gruppi animali. Comprendere questi meccanismi potrebbe aiutare non solo a spiegare l’origine della biodiversità, ma anche a proteggere ecosistemi minacciati e sviluppare nuove applicazioni biotecnologiche.
Conclusion
The study of conserved genes in butterflies demonstrates that evolution is a far more sophisticated process than previously imagined. Nature does not continuously create from scratch but instead modifies, recombines, and refines extremely ancient genetic structures.
The fact that some genes remained functional for more than 120 million years represents an extraordinary testimony to the continuity of life on Earth. Through biological wars, extinctions, and climatic transformations, certain molecular mechanisms continued shaping butterfly wings.
Butterflies thus become living symbols of the planet’s evolutionary memory. Every wing contains traces of a remote past reaching back to the age of dinosaurs.
Future research will probably reveal that similar phenomena exist in many other animal groups. Understanding these mechanisms may help explain not only the origins of biodiversity but also protect threatened ecosystems and develop new biotechnological applications.